Apomikty – rośliny, które unikają mejozy i zapłodnienia
Autorzy artykułu:
Joanna Rojek*, Edyta Ziegert
Uniwersytet Gdański, Wydział Biologii, Katedra Cytologii i Embriologii Roślin
*joanna.rojek@biol.ug.edu.pl
U roślin wyższych w toku ewolucji powstało kilka strategii prowadzących do skutecznego rozmnażania. W specjalnie wykształconych strukturach kwiatu: pylnikach i zalążkach, dochodzi do wytworzenia komórek sporogennych wchodzących w mejozę, w wyniku której powstają komórki macierzyste gametofitów, gametofity i gamety o zredukowanej liczbie chromosomów i zawartości DNA (w przypadku diploidów 2n – po mejozie uzyskany status 1n; Fig.1A). W rozmnażaniu seksualnym (płciowym) niezbędne jest podwójne zapłodnienie, aby powstały: zarodek i bielmo. Produkty powstałe po podwójnym zapłodnieniu, w postaci nasion, stanowią kolejne pokolenie warunkujące przetrwanie gatunku.
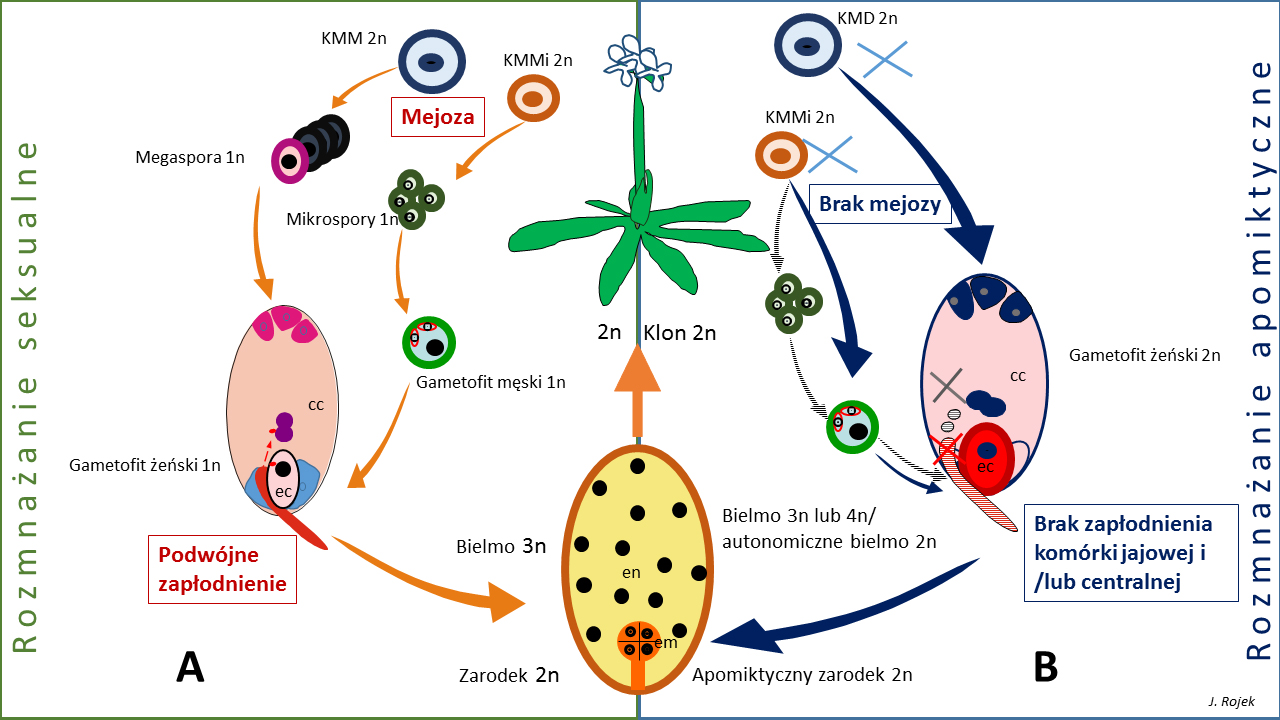
Fig. 1. Porównanie rozmnażania seksualnego i apomiktycznynego u Angiospermae na przykładzie Boechera (Brassicaceae). Zarówno w rozwoju płciowym, jak w apomiksji gametofitowej typu diplosporii, powstają komórki macierzyste spor (KKM – komórka macierzysta megaspor, KMMi – komórka macierzysta mikrospor).
A. W rozwoju płciowym, w wyniku mejozy, z jednej funkcjonalnej megaspory rozwija się gametofit żeński, który w typie monosporowym Polygonum zawiera 8 komórek, w tym dwie, ulegające zapłodnieniu: komórkę jajową (ec) i centralną (cc). Powstałe po mejozie mikrospory, rozwijają się w dojrzałe ziarna, które wytwarzając łagiewkę, dostarczają plemniki do komórek docelowych. W wyniku zapłodnienia, powstaje zygotyczny diploidalny zarodek i triploidalne bielmo.
B. W przypadku rozwoju apomiktycznego, z megasporocytów (KMD, KMMi) bezpośrednio po mitozach powstają gametofity, bez fazy mejozy (x). Mejoza może zachodzić prawidłowo w archesporze męskim (KMMi) i produkować zredukowane mikrospory i gametofity (strzałki paskowane). W przypadku autonomicznej apomiksji, brak w ogóle zapłodnienia – zarówno zarodek jak i bielmo powstają bez udziału genomu męskiego. W przypadku pseudogamii, partenogenetyczny zarodek jest odżywiany przez bielmo, powstałe po zapłodnieniu (paskowana łagiewka i plemniki). Takie bielmo może mieć niekiedy wyższą niż 3n ploidalność. Powstające nowe pokolenie jest diploidalne lecz jest klonem matki. Pozostałe oznaczenia: em – zarodek; en- bielmo, endosperma.
Rozmnażaniem poprzez nasiona, ale z pominięciem zarówno mejozy jak i zapłodnienia, charakteryzują się rośliny apomiktyczne. Powstające potomstwo jest klonem rośliny matecznej (Tucker i Koltunow 2009), gdyż nie zachodzi rekombinacja ani segregacja chromosomów podczas mejozy, ani też nie ma zmienności wnoszonej przez organizm męski.
Apomiksja jest obecna u ok. 400 gatunków pochodzących z 40 rodzin jedno- i dwuliściennych (Schmidt i in. 2015). Niektóre gatunki apomiktyczne to rośliny uprawne, np. mango, gatunki z rodzaju Citrus, tropikalne trawy pastewne z rodzajów Panicum, Brachiaria, Dichanthium i Pennisetum (Tucker i Koltunow 2009). Najbardziej znana jest apomiksja gametofitowa, w której powstaje gametofit wprost z niezredukowanej komórki inicjalnej archesporu (diplosporia, Fig. 2) albo z komórki somatycznej zalążka (aposporia) (np. Grimanelli i in 2001). Gametofit może w ogóle nie powstawać, a zarodki kształtują wprost z komórek somatycznych zalążka (embrionia przybyszowa; warto przyjrzeć się pestkom u cytrusów – jeśli rozdłubiemy takie nasiona, zaobserwujemy w nich od kilku do kilkudziesięciu drobnych zarodków somatycznych). Takie przybyszowe zarodki rozwijają się jednocześnie z zarodkiem zygotycznym i konkurują o pokarm zawarty w bielmie (Koltunow 1993).

Fig. 2. W rodzaju Boechera wiele gatunków to apomikty (na fotografii Boechera formosa; fot. J. Rojek)
U roślin apomiktycznych zarodek powstaje partenogenetycznie. Inaczej jest z komórką centralną, która z reguły wymaga zapłodnienia (pseudogamia), aby mogło z niej rozwijać się bielmo. Tylko u całkowicie autonomicznych apomiktów komórka centralna dzieli się niezależnie od zapłodnienia, dając apomiktyczne bielmo (Koltunow i in. 2004).
Apomiksja nie jest rozmnażaniem wegetatywnym (odtwarzanie całego organizmu z komórek wegetatywnych i brak rozmnażania poprzez nasiona). Dzięki tworzeniu nasion, apomiksja jest bliska rozmnażaniu płciowemu, a dodatkowo umożliwia uzyskanie potomstwa jednakowej i wysokiej jakości; utrzymuje wigor mieszańców z jednoczesnym zachowaniem rozmnażania przez nasiona. Stąd tak duże zainteresowanie budzą badania nad wprowadzeniem apomiktycznego rozmnażania do roślin uprawnych (np. Grimmanelli i in. 2012, Crissmani i in. 2013; Schmidt i in. 2014). Dzięki najnowszym technologiom naukowcom udaje się coraz bardziej zgłębić mechanizm naturalnej apomiksji i ustalać epigenetyczne czynniki kluczowe dla przełączania rozwoju z seksualnego na apomiktyczny (Crrismanii i in. 2013; Schmidt i in. 2013, 2015).
Zarówno rozmnażanie płciowe, jak i apomiktyczne, posiada znaczące zalety dla naturalnych populacji, jak i w zastosowaniach rolniczych. Rozmnażanie płciowe prowadzi do uzyskania potomstwa genetycznie i fenotypowo zróżnicowanego, co pozwala na adaptację osobników danego gatunku do zmiennych warunków środowiska i uprawę nowych, wyselekcjonowanych odmian. Przeciwnie, apomiksja wytwarza pokolenie klonalne, które jest genetycznie identyczne z rośliną mateczną, tak więc stabilizują kompleks genotypowy. Mimo, że apomiksja jest rzadka wśród roślin uprawnych, inżynieria apomiktycznych upraw jest niezwykle obiecująca i stanowić może olbrzymi potencjał i ekonomiczną wartość dla zwiększenia produkcji plonów oraz wykorzystana w innych dziedzinach rolnictwa (Spillane i in. 2005; Schmidt i in 2015 i cytowana tam literatura).
Literatura:
Bicknell RA and Koltunow AM (2004) Understanding apomixis: recent advances and remaining conundrums. Plant Cell. 16 Suppl. 1:S228-S245.
Crismani W, Girard C and Mercier R (2013) Tinkering with meiosis. J. Exp. Bot. 64:55-65.
Grimanelli D, Leblanc O, Perotti E and Grossniklaus U (2001) Developmental genetics of gametophytic apomixis. Trends Genet. 17:597-604.
Koltunow AM (1993) Apomixis: embryo sacs and embryos formed without meiosis or fertilization in ovules. Plant Cell. 5:1425-1437.
Schmidt A, Wöhrmann HJP, Raissig MT, et al. (2013)The Polycomb group protein MEDEA and the DNA methyltransferase MET1 interact to repress autonomous endosperm development in Arabidopsis. Plant J. 73(5):776-787. doi:10.1111/tpj.12070.
Schmidt A, Schmid MW and Grossniklaus U (2015) Plant germline formation: common concepts and developmental flexibility in sexual and asexual reproduction. Development. 142:229-241. doi:10.1242/dev.102103
Spillane, C., Curtis, M. D. and Grossniklaus, U. (2004). Apomixis technology development-virgin births in farmers’ fields? Nat. Biotechnol. 22:687-691.
Tucker MR, Koltunow AMG (2009) Sexual and asexual (apomictic) seed development in flowering plants: Molecular, morphological and evolutionary relationships. Funct Plant Biol. 36(6):490-504. doi:10.1071/FP09078.